Olivier Husson 3/7 : Redox-pH et fonctionnement des plantes et de la vigne

Captation réalisée dans le cadre d’une formation d’Olivier Husson en février 2022 à Châteauneuf-du-Pape.
Captation et montage vidéo réalisé par : Charlotte Brunier et Romain Mercieux.
Découvrez sur la Chaine Youtube l'Alternative un condensé de la formation d'Olivier Husson !
Conférence d’Olivier Husson : https://youtu.be/pq_Yua7Fnas
Lien de la chaîne : https://www.youtube.com/channel/UCYDa7tXvpKOTB2XxnqaaqvQ
Transcriptions
carbure de silicium du produit du co2
dans des nanoparticules
donc il pourrait bien y avoir cet effet
par la production d'hydrogène donc
réduction forte
sur la silice il ya plein de choses il
ya plein de choses à faire et biochar
plus si lisa fin de chars riche en
silice donc on
l'épée les tees et ou machin parce que
sinon je veux dire s'il faut pas qu'ils
aient du mistral parce que sinon tu les
mecs à marseille le shop
tellement plus virulent que
du biochar dans l'eau pour le pulvériser
sang sur les mains dans les poches
avec
ses batteries comme moi le plus simple
c'est de
le compresser de faire des pelles être
qu'enfin des trucs comme ça parce que
sinon dans dans l'eau mon avis tu peux
préparer quelques heures pour nettoyer
tpu levée si tu
un système d'arrosage vois ça doit quand
même pouvoir colmater mais bon pouvoir
truc d'épandage en tuellement m way
d'ottawa éventuellement comme ça mais en
tout cas voilà là des trucs en paille
voilà après si vous faites des biochar
riche en silice il faut pas monter trop
température
700 750 maxi maxi parce qu'au dessus
vous faites de la cristobalite hélas
cristobalite
c'est niveau de toxicité c'est comme
l'amiante
voilà c'est c'est bon à savoir
mais il faut il faut plutôt viser 700
surtout pas dépasser 750 degrés parce
que tu
tu fais un truc hyper cancérigènes moyen
voit donc
comment que ça se passe t-il dans les
plantes donc ça c'est comment mesure
avec les appareils les machins vous
voyez comme ça peut être galère il faut
rouler la feuille on l'à pic sur les
électrodes on verra ça mais bon c c'est
pas ça qui est pas comme ça qu'on va
avancer quoi ça c'est bien pour le
c'était très bien de pouvoir le faire
pour la recherche mais c'est sa l'étape
d'après c'est déjà ça c'est déjà
obsolète ça nous a permis de bien avancé
mais il faut que ça devienne obsolète
très vite quoi voilà donc une baisse
ignorer dogues se disait jusqu'au enfin
jusqu'aux années 2000 c'était tout
régulé par le ph en physiologie et des
depuis le début des années 2000 on
avance sur ce qui est régulée donc ça
c'était déjà un certain nombre d'années
meilleure régulation des enzymes les
pompe à protons les canaux ioniques
l'absorption des éléments nutritifs et
régulateurs de croissance la mort
cellulaire programmée l'ouverture des
stomates pouvez me parler à peu près
n'importe quoi en physiologie je vous
trouve des publics sur les régulations
redox
donc c'est il y en a de partout vraiment
de partout donc on peut
comme on est passé au travers
la perception de l'environnement dans ce
que je disais les stress biotiques et
abiotiques tout ça ça transcrit en
signaux redox et des dérégulations
les sikhs phénologique les interactions
que les hormones les dormants
germination on va passer parce que il y
en a de partout mais vraiment partout
donc voilà on sait pas ce qui n'est pas
régulé par le paradoxe maintenant et
puis pour montrer un peu une idée en
illustrant ces signaux électriques on en
retrouve un peu partout avec en
particulier des flux de calcium donc
c'est c1 lyon donc on va regarder
voilà la symphonie qui bouffe et ça
envoie un signal un fût de calcium au
reste de la plante donc là on imite avec
du cloud sur du glutamate on met sur la
plante et ça envoie un signal électrique
tout le reste de la plante là c'est une
chaîne i qu'à manger à l'autre bout et
paf
il ya un flux est un signal électrique
qui passe quoi on en retrouve donc là
s'est accélérée un mais voilà hop une
chimie qui bouffe et ça fait ça
non c'était bien accéléré quand même
ah oui ça va oui c'est oui ça fait quand
même un système nerveux pas ils peuvent
faire bouger dans tous les sens mais
c'est oui c'est pas c'est pas c'est pas
ça prend pas deux jours oui oui c'est ça
va vite voilà donc c'est pour vous
donner une idée de ces signaux
électriques et en gros ben plus
l'environnement fluctue rapidement ça va
plus ça stresse les organismes
et puis il ya des niveaux de tolérance
pareil pour tout le monde on peut y à la
même chose sur la température aussi à
papa tout supporter
et donc si une régulation qu'est
premier truc je vous disais des tampons
et doc mais c'est hyper complexe donc je
vous parlais d'hydroquinone c'en est un
des principaux mais les hormones ça va
être beaucoup lié aussi enfin ça va
jouer ça va réguler les ducks ça va être
régulée par le red dot il y en a à peu
près tous les niveaux c'est des cascades
des électrons protons enchaîne jeu mais
transfert de l'un à l'autre et puis je
perds où je gagne de l'énergie chaque
passager je récupère l'énergie où je
perds de l'autre côté en face et des
échanges d'électrons cdd rouret ducks et
puis si la capacité tampon a dépassé 20
c je vous disais de la réponse par de la
transcription formation protéines
on active des jeunes et qui font ces
gens là
donc ça leur a fallu mieux mettre dans
l'autre sens ce schéma mais c'est une
publication donc les trois gros gros
régulateurs et docks donc le plus réduit
sénat des ph n à des pc ce qui sort la
photosynthèse ça fait du glutathion
réduit glutathion oxydé qui fait de la
cid de la vitamine c est réduite
vitamine c oxydé équivalent à la
vitamine c elle réduit l'oxygéner en eau
en occident et elle va être rechargée
par l'habitation qui rechargeait par
l'un des pillages
et c'est que tout que je me passe je me
passe ça comme ça donc les gros gros
régulateur c'est ça c'est les 3 et trois
principes on va dire dans la plante dont
quoi céder roues qui tu veux mais change
des choses alors après on retrouve les
niveaux redox on voit le pourcentage de
réduit par rapport à oxyder duân adp
donc ça c'est l'oxydé ça c'est le réduit
en fonction du niveau redox le
glutathion il est là l'acidose comme la
vitamine c elle est là voilà donc on
retrouve en regardant en fait les
niveaux le ratio oxydé sur réduit ça
nous donne le moyen de mesurer le
potentiel redox quelque part c'est comme
ça qu'ils font plutôt physiologie
mais c'est vraiment ces trois là qui
sont au centre de des grosses régulation
voilà alors ce que je vous disais c'est
que ces compartimentée donc les cellules
elles fonctionnent à ph et redbox donné
la mitochondrie c est centrale
énergétique ça va fonctionner autour du
niveau du cytochrome ccc
je suis plus de tête c'est autour de 310
livres je sais moi je pense par là le
noyau est plus réduit le cytoplasme est
plus réduit par dont le noyau est plus
oxydé le cytoplasme aussi leurs
monoplaces piqué puis c'est l'espace
extra cellulaire qu'elle plus oxydé la
cellule elbaz armes dehors tous qu'il
est tout ce qu'il attaque trop
et
le noyau la transcription de l'adn ça
fonctionne uniquement à barrer ducks
alors en particulier il ya des processus
on verra sur les plans d'eau moi je peux
vous en parler tout de suite c'est
il y a
quand un virus attaque la plante arrive
à reconnaître par exemple sur le russe
tamusik jaune du riz le virus de la
mosaïque jaune dur il a besoin d'un ph
élevé pour être mobile dans la plante et
puis surtout pour se gonfler sortir la
rn et pouvoir se multiplier donc déjà il
a besoin d'un ph élevé et puis au niveau
redox en fait
le virus il va se faire identifier par
la plante général qui va produire une
protéine
pr1 protéine responsable
qui qui va rentrer dans le noyau
c'est un signal pour le noyau pour
fabriquer des protéines de défense et la
protège la paix r17 protéines de signal
quand elle est réduite elle est sous
forme monomère donc séparés et quand ces
oxydes et elle se polymérise ça devient
trop gros et le rendre plus dans le
noyau donc en fait le virus ils bloquent
le il a besoin dans redbox élevées pour
que cette peine soit oxydé pour pâques
la pente puisse se défendre
et il ya plein de processus dérégulation
qui vont être comme ça de près
transcription ça c'est avant la
transcription et puis après quand il ya
eu des enzymes toutes les toutes les
protéines qui sont souffre et en
particulier quand elles sont réduites il
ya quand il ya deux ans il ya des achats
crochet os
et puis quand elles sont donc elles sont
fonctionnels et puis quand ça s'oxyde
2h parts il ya les 2es qui s'accrochent
charpy la protéine qui ne marche plus
donc ça peut bloquer avant la
transcription ou après la transcription
et en particulier ces protéines avec de
soufre et de s la basse et des switchs
reddox c'est à dire que c'est réversible
sa source ac en fonction du niveau redox
ça va se replier ou ceux
de se déplier où se refermer donc il ya
vraiment niveau physio des processus y
en a un paquet aussi qu'ils sont connus
comme ça voilà donc mais c'est très
compartimentée aurait alors que leur
place - six ans - 900 millivolts quoi
c'est c'est très réduit en fonction des
côtés du qu'il a co id enfin il ya des
choses donc ce que je vous disais c'est
que les micro organismes qui régulent
leur environnement avant tout
c'est par les populations donc ces
biofilms qui vont modifier complètement
les conditions ph red hot qui vont
favoriser
leur développement en interne ça quand
même pas de grosse grosse capacité à
réguler donc s'ils sont tous seuls
isolés ils en arrivent à faire grand
chose pour soi qui a besoin d'un certain
nom d'un certain niveau de la population
pour arriver à réguler
et donc ces micro-organismes ils ont une
grosse capacité à modifier le milieu
enfin
ajuster le milieu à leur niveau
favorable donc les plantes elles vont
utiliser ça pour réguler donc
ça va c'est très spécifique ça va
orienter une plante carencée en
manganèse elle va faire des exodes a qui
vont favoriser des micro-organismes qui
solubiliser le manganèse donc c'est
c'est très très spécifiques les plantes
elles régulent le milieu interne et le
milieu externe donc au niveau de la
rhizosphère
est ce qu'il faut comprendre c'est que
c'est pas je régule plus ou moins bien
et je fonctionne plus ou moins bien
c'est doit réguler à tel niveau ça me
coûte plus ou moins cher j'ai de quoi me
payer ou j'ai pas de quoi me le payer
quoi
c'est pas une dérive
la cellule qui ne fonctionne de moins en
moins bien c'est vraiment j'arrive plus
à ces sales histoires de coups de
pouvoir se payer les animaux bas c'est
comme les comme les plantes c'est en
réguler le milieu interne et milieu
externe on entretient des populations
qui nous on essaye de favorables et les
plantes elle se comporte comme ces
élites elles et les conditions extrêmes
de ph ou dreadlocks donc elle modifie
directement dans l'eau au niveau de la
rhizosphère pour les exsudats et via les
micro organismes qui les orientent ça
peut aller
deux unités ph pour corriger le milieu
on a même vu c'est même plus en fonction
de la zot qui a absorbé la forme d'un
vote
en gros ça va me baisser en milieu
alcalin ça ramène toujours vers neutre à
peu près et en milieu acide ça va le
call in iz et enfin ça va ça ramène
proche d'une ode légèrement acide on y
retourne là autour de la racine ça on le
retrouve au nez au niveau de différents
de l'a7 péage sur quoi on est sûr
c'est du maïs de 40 jours ça ramène
toujours le ph autour des mêmes niveaux
au niveau du remdoc c'est pareil là on a
les potentiels redox on à la distance
par rapport à la racine en mm donc là on
est en rizières sur un truc totalement
asphyxiants et bain juste au niveau de
la racine ça a réussi à remonter 2 -
presque - 300 ha ha 50 millivolts
s'est pas encore suffisant normands
voient que le riz a réussi à faire
quelque chose juste autour sa racine sur
un truc qui a moins de sens millivolts
le sol ça joue sur sur 2 mm et ça
remonte et puis sur un sol qui est à
cent mille vols donc son millivolts
c'est quand même bien naze fiction déjà
c'est on est bas et ben ça joue sur 3 4
mm et sa remontée à chaque fois ça
remonte autour de 400 mille volts
et dans l'autre sens ça c'est un autre
type de manip c'est de la féverole sur
un seul oxydé on met une électrode et
puis on regarde pousser la racine quand
la racine passe devant l'électrode ça
passe de 700 mille volts à 380 millions
dhs
pourquoi donc la racine autour de la
racine elle maintient au taux de 400
mille volts ph légèrement acide
donc c'est alors à des différences entre
l'apex les autres partis racines mais en
gros la plante elle corrige le milieu et
ça ça nous coûte de l'énergie du milieu
est déséquilibré plus l écoute de
l'énergie
pas plus compliqué que ça alors il ya un
effet direct des exumas et puis il ya un
effet indirect par la microflore j'dis
est spécifique mais globalement les
micro-organismes qui consomment de
l'oxygène qui ont ensuite été gazés donc
c'est donc ils réduisent le milieu
et au niveau du ph en général salle
baisse mais c'est souvent neutralisé
dans l'essonne
voilà alors après quand on regarde les
publications
les pourcentages de de la photosynthèse
des produits photosynthèse qui sont
descendus au niveau des racines la
raison des positions donc sous
différentes formes entre les
les exsudats racinaire les cellules qui
meurent des racines ces trucs là bas
dans les publie sa va de 5 à 80 % de la
production photosynthétiques
donc derrière c'est pas bien dur de
faire une hypothèse assez simple c'est
que plus on les déséquilibres et plus ça
consomme
et que 5% c'est qu'on ne sait bien
équilibrée que la plante elle sous
traite juste la prospection et qu'elle
n'a pas besoin de grand-chose
80% c'est souvent dans des milieux super
déséquilibrer les toundras étaient les
choses comme ça non plus ça produit pas
énormément donc
on assure leur et docks sont naïma loire
sur le ph on voit ça on voit que
l'exultation racinaire augmente quand
pulpe et hachez déséquilibre
est donc ça c'est une public dont je
vous parlais tout à l'heure c'est du blé
qui pousse à ph 4 5 et demie 7 et 8 et
demi donc là on est sûr de les solutions
nutritives ces débits de silice et
pillant nourrit
corrige le milieu ça nous donne le
niveau de la rhizosphère aussi en même
temps la ccc traite en poney et donc si
on regarde le pois frais si on met 100 %
à ph est ma perche cet ennemi on perd
5,5 et demi on perd 5% on perd 10 7% à
ph 8 et demi on perd 12% à pêche 4
donc ça c'est sûr le pois pois frais en
pois secs c'est un peu près pareil enfin
voilà on perd 6 7% donc ces gros vite
ennemi c'est aussi déséquilibrée que 4
puis quand on commence à regarder les
racines mon père 6% à péage 5 ennemis
32% à ph8 ennemis 35% à ph 4
c'est
alors c'est du bluff c'est pas de la
vigne mais
il y avait quelque chose qui se passe
quand même qu'il en va pas très bien et
puis au relais longueur on va passer et
puisqu'il y est intéressant c'est quand
on regarde les niveaux d'oxydation des
plantes donc en fonction du ph
la solution là où poussent les racines
donc ça c'est des marqueurs de stress
oxydatif donc mt à lhassa celle oxygéné
ça c'est un autre type lors l'activité
d'une enzyme qui lie détoxifie donc ce
qu'on voit et ben
c'est très oxydé c'est aussi oxydes rph
huit ennemis kph 4 et puis à pêche
chaque ennemi ces intermédiaires par
rapport à ph est on le retrouve pour
tous ses marqueurs là si on regarde
la chlorophylle sait danser dans l'autre
sens donc un gros passage ou soit
photosynthèse et puis si on regarde le
ratio vitamine c réduite sur vitamine c
oxydé au glutathion réduit sur plus
tatillons oxydé si les deux gros
régulateur
on voit bien que c'est pareil voilà il
ya peu de ya peu de 8 est enfin leur
assise beaucoup de ces beaucoup plus
oxydé / ph 4 et 8 enfin pareil ph 4 et 8
ennemis et par rapport à ph est c'est
fait c'est poussé plus de trois fois
c'est très oxydé donc on voit l'impact
du péage sur l'oxydé d'une perche
puissance sur l'oxydation de la plante
alors après à comprendre c'est que les
feuilles où il ya la photosynthèse ça
c'est pas bien dur à comprendre c'est
celle qui sont plus réduites sur le plus
acide
ça ça se lit à travers ses ratios
vitamine c est réduite sur vitamine c
oxydé ou glutathion laëticia va être
intermédiaire et puis il va falloir
qu'on rentre dans le détail parce que
les différents types de pathogènes ils
sont à différents ils vont ils furent
pas n'importe où dans la plante quoi
donc il va falloir comprendre si c'est
sur leurs racines sur les feuilles ou
sur les fruits ou les fleurs si c'est
dans le milieu extracellulaire dans le
dilemme ou dans le flou m
donc le flou m c'est super c'est très
alcalin je vous ai expliqué pourquoi ses
dépens donc c'est donc ph cet ennemi à
huit ennemis général
c'est pour maintenir un gradient de
protons qui permet de charger c'est ce
flou et en sucre caisse et les ponts de
sucre aux protons alors c'est pas un
proton qui tire un sucre mais c'est
pareil des roues il ya besoin de protons
qui passe pour que le pour que le sucre
pasqua donc on comprend vite que si
si on monte à huit et demi en ph sur le
sol ça ça n'a pas chargé pareil coin et
donc ça c'est très tamponné et il ya une
conductivité électrique élevée aussi si
je très chargées en sucre conductivité
électrique élevée c'est pour remplacer
les virus après ils sont en général à
conductivité électrique élevée
le dilemme il est beaucoup moins
tamponné et il a ph plus acide et puis
dépend beaucoup du sol
ce qui est dans ce cas dans la solution
du sol des conditions milieu extérieur
exil et là j'avoue
j'ai pu travailler en ligne je suis papa
tôt
sur la ville mais ça devient assez
simple on comprend vite après
et puis milieu extracellulaire lui il
est très peu tamponné et
al'inverse des cellules là les cellules
elles sont très très tamponné sitôt
place cph 7,3 à peu près toutes les
cellules donc et c'est
voilà alors après le xylem bas en
fonction de comment on l'extrait on va
avoir des niveaux ph différents donc
c'est ça c'est pas si simple que ça non
plus d'aller dire on va mesurer dans le
dilemme dans le flou règne un truc faute
en fonction du mode d'extraction les
méchants ils ont ils ont c'est ph là où
ceux là où cela quoi donc il faut pour
ça que je passe pas trop de temps ici il
mesuré aussi précisément que on les
publics et puis on voit alors après les
gradients ph rédac ce dans la dans la
plante alors j'ai beaucoup travaillé sur
le riz donc qui va d'ailleurs pas mal de
trucs sur le riz je vais vous dire qu'on
fait sur la sur la vigne
déjà la base de la feuille c'est un ph
plus élevé à red box plus élevé donc ap
opus ph le plus élevé par rapport à la
pointe de la sphère
des champignons qui se mettent
alors ça c'est pour le rire sur les
feuilles de vigne il faut voir la
feuille de riz ça s'explique assez
facilement c'est que l'ailé comme ça et
donc là il ya peu de peu de lumière et
là il ya le maxi de lumière la
lapidation tourner
ça aussi mais en tout cas c'est c'est
beaucoup lié à la photosynthèse donc la
partie de la feuille de riz qui à
l'horizontale au dessus elle capte plus
de lumière que la partie qui combat
après peut-être des transferts aussi qui
se font mais savoir ce qui est
intéressant de là aussi de commencer à
avoir c'est que là en fait en bleu c'est
du riquet cultiver les pieds dans l'eau
c'est ça qui est intéressant avec le riz
c'est qu'on peut lui mettre les pieds
dans l'eau donc lui mettre un red sox du
soldat et puis
lui s'en tire quand même si on fait ça
du maïs où ça va pas le faire longtemps
quoi va pas faire pas cultivé pendant
deux mois et anvers et c'est en
pluviales donc des conditions bien plus
oxydé et puis c'est à 40 jours et à 60
jours leclerc à 40 jours le fond c'est à
60 jours donc on voit qu'il ya des
différences de niveaux de dreadlocks 6 6
2 ph donc le leur et qui pousse les
pieds dans l'eau donc sur un milieu
réduit plutôt alcalin il est plus réduit
et plus alcalin si la voile et le riz
qui pousse sampan pluviales il est
celle inverse quoi donc c'est assez
logique et un impact du milieu sur la
plante
et puis en 40 jours et 60 jours on le
verra mieux après donc voilà
donc ensuite si on prend un plan de ris
qu'on éclate on a le ph en rouge le leur
et docks sang bleu et le red rocks
corrigea ph est en verre c'est comme le
pe plus perche
donc ce qu'on voit en fait c'est
qu'alors le développement d'un plan de
ris à la première talent qui sort mais
en gros ça veut dire quoi ça veut dire
que cette feuille la cette feuille là et
cette feuille là ont le même âge
celle là celle là est celle là ont le
même âge voilà une puis en descendant
voilà ça les plans bas sont plus
vieilles on a les plus jeunes donc ce
qu'on voit c'est que la feuille en eau
est toujours plus alcalines que les
feuilles plus bas donc alors je veux pas
dire toujours dans ces cas là on en a 6
ans le retrouve on a fait pas mal de
fois donc on voit et puis le péage soit
là on est à 6 91 6 88 6 93 ouvriers on
est très très proche là là on est à 6 84
6 82 et
6,87 et on est sur des ccc une
demi-heure quatre répétitions à chaque
fois lassé moyenne donc voilà on a
vraiment un ph
la jeune femme y est plus alcalines et
les feuilles en descendant elle ceci dit
fille
au niveau du red box est bien là jeunes
feuilles elle est de 265 263 264 c'est
dur de voir précisions mieux presser
haut et ça c'est joli cd les répétitions
statistiques derrière ça c'est
l'huissier publié là dedans si vous
voulez la jeune fille en fait elle est
bien plus oxydé que la deuxième ou le
troisième
jeune fille elle est en train
de développer la photosynthèse n'a pas
sa capacité de photosynthèse complète
encore donc sûr de la vigne en fait on
retrouve ça pas sûr juste de trois
feuilles on retrouve ça sur jusqu'à 8 9e
donc ça veut dire leur et docks s'ils
baissent de la toute jeune feuilles
jusqu'à la
hervy terre-neuve parle et puis après
sur la ligne c'est relativement stable
jusqu'à ce qu'on arrive à la feuille en
face de la grappe
ou la leur et doc se remonte à priori
c'est parce que ça commence à transférer
assez logique quoi mais après pour
raisonner l'apparition des maladies de
manière différentiel en fonction des
feuilles et ces choses là ça va être
vraiment important
voilà donc la deuxième et la troisième
feuille
qui sont pleine activité
photosynthétique son red sox minimum et
puis là et puis après on vieillissant
basse et pluvieux c'est la photosynthèse
marche moins bien et puis il ya moins de
lumière on commence à être sous les
autres
donc le red dog se remonte et puis donc
les champignons il en est souvent par là
et puis on les voit beaucoup là
donc oil en gros
c'est très très lié à la photosynthèse
donc nous sauverait quand on mesure on
mesure le tiers du milieu de la feuille
en pleine plus jeunes feuilles en pleine
activité photosynthétique
pour pouvoir comparer
dvd plante si si c'est une plante vous
prenez la et puis l'autre vous prenez là
bas c'est à dire c'est pas pareil mais
en fait c'est pas pareil parce que vous
n'avez pas pris au même endroit
d'ailleurs je sais pas si vous
connaissez les analyses de selles
quelqu'un
l'analyse de scène en gros on prend
cette feuille là et puis celle là
pour pouvoir justement à voir les deux
extrêmes qua
non
ça c'est bien expliqué meurt sur la
vigne je sais pas lequel il préconise
mais en tout cas en général c'est une
moi je moi
il on préconise certaine nous passons la
vie n'y est pas en général ils disent
une jeune et une vieille mais
voilà alors cette ce gradient là on le
retrouve sur le maïs sur le cotonnier
sur le niébé enfin toutes les plantes
comme qu'elle en a on à dépiauter tous
les plans et on a fait des mesures et
des mesures des mesures
voilà ce que je vous disais sur la vigne
en bois on m'avait fait savait que les
cv la
1ere demi
pas loin de chez moi jeunesse donc son
on retrouve voilà jusqu'à la 89e le raid
hawke's bay c'est après il est assez
stable jusqu'à la feuille en face de la
grappe
voilà près d'un fonds par les
légumineuses par exemple qu'à l'open yom
c'est une plante ouverture en tropicale
il est là l amo kuna elle est là vous
voyez qu'on a on a 200 milles d'écart et
de phd car
c'est de légumineuses
à feuilles larges de même milieu
donc c'est vraiment des traits des
plantes le stylo dantès lui il est là
c'est surprenant ce maïs est pas si
surprenant que ça s'ils osent entes elle
doit faire tourner c'est ce qu'on
appelle la luzerne tropicale c'est une
plante de couverture assez magique qui
fait des trucs super ça refait les
soldats on a des plans super sain
derrière le riz en fait évidemment elle
fait beaucoup tourner ce qu'elle est et
oxydé mais vous voyez donc laissez pas
légumineuses graminées et c'est
c'est même pas espèces ou un trèfle
blanc trèfle violet complètement
différent donc cédé très d'utilisation
de la
l'énergie alors là pour comprendre comme
s'il ya la photosynthèse et ben on se
dit que dans la journée ça va bouger
donc on s'est tapé des mesures c'est à
cotonou 6 heures du matin à six heures
du soir et on est sur l'équateur 6e le
jaune la selas et la lumière était
mesurée à chaque fois conseiller mesure
on a une mesure de la radiation solaire
donc 6 heures du matin c'est avant le
lever du soleil et puis six heures du
soir c'est le coucher du soleil
donc les trois couleurs ces trois
variétés de pury là on est à 40 jours
après semis donc le potentiel redox on
voit battre le matin sa chute et puis ça
se stabilise et puis ça remonte sur la
fin c'est c'est assez logique toute la
nuit a pas eu de photosynthèse
donc donc la plante et plus oxydé puis
la photosynthèse sonnant marshall a
réduit et au passage balle et pas mal de
maladie il se développe surtout le matin
pour le ph on a une légère baisse
défendre un peu pareil mais c'est pas
forcément la sio là c'est pas très net
il ya des fois c'était beaucoup plus
nette et puis si on regarde vingt jours
après
je fais ces deux fois voilà si on
regarde 20 jours après et ben on a
toujours cette forme
logique
auxi dès le matin puis ça serait aussi
le soir le ph là c'est plus net qui
baisse puis qui va remonter
ce qu'est ce qui est marqué c'est que la
déjà la fenêtre pour mesurer il faut
mesurer
entre 11 heures et 4 heures après
peut-être qu'en plein cagnard attentif
et 40 degrés ici ça se referme un peu il
faut ensuite ça mais en tout cas
cessé
si on mesure
ça nous arrivait dans le bordelais qu'on
mesure 5
cinq plants d'une parcelle le matin à 9
heures on passe une heure et demie à
mesurer cinq plants d'une parcelle et
puis qu'on va dans une autre on a fait
autre chose et puis quand mesure 5
plantes en fait on se dit bah tiens la
première parcelle est bien plus oxydé
voyez vite où je veux en venir c'est pas
que la première phase avait plus oxydé
c'est que c'est qu'on l'a pas fait au
même moment ils s'étaient pas stable à
ce moment là donc au moins se met sur la
zone à peu près stable mais l'idéal
c'est de faire une feuille d'une
parcelle une feuille de l'autre une
fuite une feuille de l'autre et de
revenir là ça évite ce genre de problème
c'est plus compliqué par contre une fois
qu'on aura ça
vous allez faire 10 feuilles savons
prendre maxi 10 minutes en marchant elle
redit minutes de l'autre côté voilà
c'est réglé voyez le nombre de problèmes
que ça va débloquer le truc aussi ce
qu'est marqué donc c'est que le red sox
des plans qu'il est bien plus bas que le
red box du sol les seuls on parlait de
400 mille volts là on est à 200 et 250
donc ça c'est heureusement sinon
système n'y aurait pas d'énergie qu'il
rentrerait dedans et puis la
photosynthèse et de la réduction de
l'acidification on a vu et le
vieillissement la gemmi si oxydation
acidification
et c'était d'études au bénin sur le riz
on l'a fait ça on l'a fait quinze fois
mais bon c'est bien s'est bien amusé une
seule fois où on n'a pas eu cette courbe
qui descendait et puis qui remontaient
c'est qu'on avait un truc qui était tout
plat c'est on a mesuré un lundi après un
week-end où il ya eu deux jours non-stop
d'orage
orage ô rage ô rage ô rêve donc on se
dit que
il doit y avoir quelque chose là
derrière l'oral je dois recharger en
électrons pas mal et
donc c'est pour ça que souvent les
plantes sont superbes est la preuve
après une pluie d'orage si la pluie de
grêle
un bémol
mais donc voilà il ya quelque chose à
fouiller la dernière aussi entre une
pluie d'orage une pluie normale et de
l'eau du robinet on n'a pas les mêmes
redox
et votre chien il va pas boire la même
laissé le choix
il va sur la pluie d'orage
je me suis amusé à le faire
voilà alors je me dis je disais voilà
c'est vieillissement oxydation
acidification et
toutes les mesures qu'on a fait sur le
riz en côte d'ivoire au bénin on en a
fait des dizaines de milliers en
vieillissant on avait toujours ça on
fait les mesures sur le riz en france et
on se retrouve avec une alcalines
isation en vieillissant
et donc grosse question leur premier
truc on dit c'est en camargue au bénin
on est sur des ph un quai en camargue on
est à péage cet ennemi donc mans dans
une logique et puis on est sur des pots
en serre et on a la même chose la
localisation et sur des terreaux à
quatre et demi
alors c'est peut-être la flotte on est
on sait peut-être l'eau dedans et
peut-être par qu'on transfère ses des
terreaux fait spécialement avec da puzzo
la note est réparé à magnétique on a des
plans super revers magnifique et
effectivement c'est 8 euros à pêche
quatre et demi au départ et arrosé de
l'eau plutôt à cet ennemi 1,8 donc on se
dit est ce qu'on n'est pas en train de
faire une pile avec sa
fille la proton quelque part donc c'est
à fouiller ou est ce que c'est des
histoires de l'attitude de lumière de
choses comme ça mais vous voyez on se
pose plus qu plus on fait des mesures
pour une pose des questions quoi
là dans les serres là c'était moi
c'était fait au mois de printemps était
donc c'était essentiellement de la
nature ayant
je vous l'avais même pas de l'ambiance
et
c'est là où j'ai découvert ça
allait augmenter la porosité ouais enfin
c'est eric c'est parce qu'elle est très
légère baisse alors pour bath a réservé
2
le day 2 3
c
voilà alors après quand on travaille sur
la pathologie je suis pas sûr que c'est
une bonne idée de faire des terreaux
super bon comme ça
on va pouvoir s'amuser en recherche à
créer des milieux qui favorise les
maladies sur lesquels on veut travailler
quand j'étais à madagascar j'habitais
juste à côté desserts de quarantaine
pour importer des variétés de riz par
exemple ça passait par la quarantaine
dont heureusement j'ai vite et juste à
côté parce qu'on en a à faire entrer
beaucoup et donc qu'il prenait il
prenait les semences qu'on en est et on
payait des dépôts de terreau et cultivés
en serre fermé 2
des semences qu'on avait importé pour
voir s'il y avait des en particulier la
pellicule arioso ce champignon sur le
riz qui se développait sauf qu'ils
mettaient des bon terreau en irrigué
donc en fait il le mettait dans des
conditions qui sont pas du tout
favorable ce champignon
est donc après pour travailler soit sur
la sélectionner si on veut sélectionner
des variétés
et qu'on veut les sélectionnés sur la
résistance à des maladies et qu'on les
met sur un milieu qui favorise pas ces
maladies on va présélectionnés grand
chose quoi et sur les variétés de riz
dans les publications les mêmes variétés
sont classés complètement différemment
en fonction des publics je pense que le
lait raison c'est que c'est que quand
elles sont sur un milieu qui est plutôt
équilibré devrait être plutôt résistante
on sélectionne pas grand chose quoi
voilà et donc après sur le red sox
corrigé à ph est
voilà ce que ça donne c'est qu'à 40
jours on est
on a vraiment des grosses différences
entre variété
et la juste polis truc direct c'est que
la variété rouge c'est une variété très
sensible à la pellicule are you c'est ce
qu'on utilise dans les combos
manifestante quand on veut justement
sélectionné sur les périls arioso pour
être sûr qu'il ya des sports partout on
met ça régulièrement parce que ça va se
multiplier à fond et que ça va le savent
a dit m cette variété là elle est très
résistant
donc là on parle de
30 mille volts 60 jours la floraison un
truc c'est les feuilles et tout c'est en
tout est en train de se vider pour aller
sur
sur la fleur est en train de transférer
l'énergie vers les fleurs
donc quand on est dans de bonnes
conditions peu plus punchy baisse au
moins jusqu'à la floraison quand on
mesure la feuille et dont je parlais
voilà mais en gros le résumé c'est que
tout ce qui va modifier la photosynthèse
savoir un impact sur le ph redox qu'on a
vu la nutrition qui va qui va jouer déjà
et puis après il ya toutes les
conditions du climat quoi il n'ya pas de
soldes
la lumière est température l'eau
avec le sol qui tamponne un peu sur
l'eau s'il est bien fait voilà donc ça
je vais passer mais en gros dans la
journée la photosynthèse on va plutôt
comme ça quand on met ph et redox en
même temps la photosynthèse ça nous
amène vers le bas à gauche
on revient
alors à près sur le côté vieillissement
voilà le genre dessert on s'est amusé à
faire on fait pousser du riz dans des
pots donc on a en irrigué il y en a en
pagaille riez
et puis pour voir le vieillissement mais
on a on a planté plein de paul même jour
et puis on a mesuré tous les dix jours
et puis on a planté dépôts tous les dix
jours et on a mesuré
après 70 jours tous les pots avec des
âges différents pouvoirs si c'était que
ce soit vraiment un effet de l'âge pas
que ce soit en effet des conditions de
lumière depuis une machine qui a pu
avoir donc et on l'a fait en pot on l'a
fait en grande parcelle en côte d'ivoire
la droite à gauche et le bénin à droite
c'est la côte d'ivoire ce qu'on retrouve
c'est que au niveau du ph alors on peut
prendre un petit peu de temps là on a le
ph du sol à 0 5 05 et 5 10 sur le sol
qui conduit en pluviales d'accord donc
on est sur un sol acide à la base et qui
est oxydé et puis par contre quand on
met sous l'eau et ben obtient lé ph qui
monte là on n'a qu'à dix jours en fait
on est on a dû partir de là à peu près
puis ça monte vite en ph la submersion
fait monter le ph le céleri qui
poussaient sur le sol
sec et acide il est il est plus acide
que le riz qui pousse sur le sol qui est
plus elle cale enfin sur dans les
conditions de milieu avec la submersion
qui montre le ph donc on a un impact du
ph du sol sur le péage de la plante et
on a un impact alors sur les conditions
redox et bats la submersion ça fait
chuter complètement donc on parle plutôt
de 550 mille volts et puis ça s'écroule
et on descend à on était à cent
cinquante mille volts à la fin alors sur
les différences sur le riz ça s'écrase
parce qu'elles sont plus faibles mais on
voit bien que le riz sur le sol réduit
est plus réduit que le risque sur le sol
occident
oui
ça pose est est il qu'en y réfléchissant
je me dis que je pourrais faire aussi le
ph en fonction du raid du sol
on aurait donc en fonction du ph du sol
plutôt redox de la plante en fonction du
ph du sol ce pourrait être une bonne
idée de reprendre cette année du quiou
10 aix pour corriger le ph on fait
tourner des pompes qui oxyde faudrait
que je regarde ce que ça donne à
l'époque j'avais pas pas percuter là
dessus alors après si on met le ph et
dreadlocks
en fonction du temps à dix jours et
voilà ce qu'on voit c'est que c'est que
série la condition pluviales donc oxydé
à ces débats y sont plus acide et aux
fonctions quasiment aussi acide est
oxydé à trente ou quarante jours que
l'autre à 70 jours donc il vieillit plus
voilà avec le bémol que je vous dis
c'est que peut-être l'eau c'est
peut-être
alors après si on fait sur du c'ur du
blé la banderole et 6 ces cyber
charpentier c'est ce qu'a permis de
calibre a laissé les spectre au parti
qu'est sur le blé donc c'est d'ultras du
semis direct en bail liant au dessus
c'est duduche isole un travail un peu de
travail du sol même si c'est pas au
travail du sol on voit qu'il ya quand
même
ouais pratiquement 10 milady millivolts
d'écart entre les deux et là c'est tout
c'est cinq variétés mélanger donc après
c'est pas énorme et le lush isole c'est
du bon travail du sol quoi c'est pas
voilà yat il ya toute une gamme c'est
pas
c'est pas là c'est pas c'est pas c'est
pas du massacre il tient hubert il a
gardé ses parcelles depuis depuis 20 ans
il a gardé 24 mètres donc double mètre
12 maîtrise une rampe de pu lever en
labourd il ya tout en semis direct et
sur ces quatre systèmes il a il a 12
mètres en labourd et il arrête cette
année ça c'est fini ça il a ras-le-bol
de passer plus de temps à faire les
traitements sur le sur son hectare de
labour que sur les 50 hectares d'un côté
donc nettoyer les pu lever est passé
trois fois plus souvent donc il y
maintenant j'ai gardé ça pour la
recherche un moment maintenant
j'arrête de faire ça mais il résulte
mois on a fait sur le sur l'orénoque du
sol oui on a fait quelques pas
sur une seule ferme comme ça c'est un
peu dur on essaie justement de faire la
thèse
sur le blé qui est comme ça il ya
22 couples de parcelles semis direct
conventionnelle
sur toute la façade atlantique enfin pas
tout cas façade atlantique sur sur un
triangle
poitiers nantes tours à peu près donc là
on va avoir un ajustement tous est
suivie avec des semis direct de 3 25 ans
donc on va pouvoir une gamme certains
des animaux pas d'autres certains commis
du basale pas d'autres
on va pouvoir commencer à sortir des
choses
ça on doit pouvoir publier parce qu'on a
suffisamment de choses voilà ça c'est
sûr le ph donc au moment de la floraison
il ya un truc un
passe quelque chose voilà et donc le
leur et ducks corrigé à ph est ça donne
ça donne ça quoi donc on a le red box
plus bas et le ph plus bas mais on est
des deux côtés
bourges problème
si une plainte par exemple on arrive pas
ça peut créer aucune idée
ça pourrait vouloir dire non mais je
sais pas je n'ai pas de chez pas d'infos
là dessus
sur les fleurs on a les mesures qu'ils
aient eu dreadlocks et si mylène
souligné qu à fait sûr
sur des pétales d'abricotiers pour
regarder la moniliose
donc il faudrait voir comment ça se
passe globalement la fleur c'est donc
pas le pollen est aride hawks très bas
donc
je voudrais pourrait voir
voilà alors après donc voilà ce que je
vous dis oui mais si je vous disais
c'est de l'acidification là si on voit
que si on est plutôt en al talhi
nisation ce qu'est ce qui est marrant
aussi ses différentes variétés de riz
alors obsédée ça c'est des variétés
japonica tropicaux on à fait différents
types il ya les rires du japonais cac
est plutôt irrité il ya de l'indépendant
qui est plutôt pluviales il ya de lundi
qu'à qui est plutôt autres irriguées qui
poussent dans des conditions différentes
donc là c'est que des variétés japonica
on voit qu'elles ont des ph et des red
sox différents donc là c'est sur trois
sabres et surtout ça doit être 30 jours
45 jours et 60 jours les trois les trois
mesures qu'on veut on voit qui est des
variétés qui maintiennent pas mal alors
que déjà qu'ils sont différentes en ph
et puis on voit qu'il ya des variétés
qui modifient qui contrôle peu le ph qui
s'oxyde pas trop et des variétés qui
contrôlent plus pêcher au début puis qui
finissent par partir en
s'oxyde pourquoi pas lequel contrôle pas
quand elle met se mettre à contrôler
elle s'oxyde à fond pour pouvoir
contrôler le ph elle s'oxyde donc elles
doivent maintenir tant que ça fonctionne
à peu près au niveau du ph et le garde
et puis et puis comme ça quand ça va
plus il s'oxyde pour maintenir le ph
dans les zones où ça fonctionne quoi
là c'est pareil on voit bien c'est de
cette variété là alors là on est sur des
japonica tempéré donc on est sur des
gammes différentes mais on voit celle là
là elle contrôle très peu son ph et du
coup elle s'oxyde pas trop celle-là il
contrôle beaucoup plus le ph elle part
en flèche là aussi
donc ça au niveau des cépages est plus
porteuse règle
ah oui il ya du taf la haye mais au
moins y a les clés pour comprendre ce
qui se passe
avançant ses clés là on n'y arrive pas
alors ça c'est des résultats
préliminaires parce qu'on avec trois
mesures mais avec les scanners on a on
peut remonter en arrière quand on sait
la calibration donc ça va pouvoir
augmenter la puissance statistique parce
que maintenant qu'on a calibré on peut
avoir le ph redox à partir du scan mais
bon j'ai pas j'ai pas quoi faire les
schémas j'ai juste reçu les données
l'autre jour mais en gros
suis plus quel cépage chai ça me va bien
donc le témoin on est on est là on
traite à la vitamine c ou aux cuivres le
rouge et le vert c'est kif kif on change
pas vraiment de jaune par contre on
mélange les deux
ans oxydes pour moi ce qui se passe
c'est que le cuivre envoie un petit mais
c'est un petit stress oxydant ce qui
fait réagir la plante qui se réduit
voilà qui salent calviniste un peu
qu'ils baissent le
h
alors que la vitamine c
le réduit naturellement en apportant de
l'énergie ça c'est juste sur une mesure
c'est pour ça qu'il faut le faire dans
le temps aussi sinon on n'arrivera pas
si on mélange les deux on envoie un
signal d'oxydation avec un truc de
réduction elle sait plus où elle en est
ses antagonistes c'est de ses deux deux
traitements qui sont antagonistes pour
moi
c'est justement ça va nous permettre de
repérer ce qui est ce qui va ensemble
comment combiner des leviers de ce qui
est antagoniste qu'il faut surtout pas
mettre en même temps
et pour ça qu'il va falloir suivre ce
que je disais c'est que sur une fois je
pense que peut-être qu'on fait c'est que
la vitamine c elle réduit il y en a
exactement des électrons et que le
cuivre stresse un peu la plante qui
corrige et que si on le fait à
répétition
elle va finir brel risque de finir par
s'épuiser pas pousser à c'est parce que
ba ciel corps et elle change son
métabolisme elle fait moins de
croissance pour se réduire
voilà pour se protéger en se disant
attention ça commence à chauffer là je
me
sinon c'est pas ça dépend par rapport à
quoi c'est à priori oui on il est moins
bien pas il est plus acide
ça c'est les feuilles de la vigne ces
deux-là 8/9 feuilles enfin une feuille
en plaine en pleine activité
photosynthétique et
exposé à la lumière
c'est pareil le coût de la lumière on
n'a pas parlé mais après il y aura
l'orientation dérange
en du nord sud et de l'est ouest
ça va pas faire la même chose il y en a
encore à une phase toujours au soleil
l'autre toujours à la
bourre à l'ombre alors que dans l'autre
cas ça sera au soleil le matin
d'un côté et l'après midi de l'autre
côté on sait que ça marche moins bien
beaucoup moins bien je vous rendent pas
dans les phases de la lune
je rentre pas dans les phases de la lune
john chiem ben dilla il ya deux semaines
par mois où ça marche moyen il ya une
semaine où ça marche super et une
semaine où ça marche pas
john keynes en américain qui travaille
beaucoup sur les analyses de ce mec est
que sur l'agriculture de régénération
cocaïne et une histoire intéressante qui
fait des choses super intéressantes
il est sur des dizaines de milliers
d'hectares aux états unis sur plein
plein de culture travaille beaucoup et
qu'ils analysent de cette fois il fait
des plans de plein de webinaires super
intéressant c'est de l'anglais simple il
parle il parle lentement c'est pas c'est
pas le texan
incompréhensible
puis on peut même mettre la traduction
par contre vous ne verrez pas sa tête
c'est un ami
donc il se fait pas il sera filmé
mais en gros vol quand il avait 14 15
ans son père était un des gros
distributeurs de produits phyto et il
lui c'est les essais dans la ferme
depuis un moment ils ont récupéré une
prairie à côté d'une parcelle de melon
épuisant fait du melon et puis et il est
remonté l'allée rencontré plein de plein
de chercheurs plein de monde partout et
s'est construit le modèle puis l'a fait
une boîte de conseil et puis suivront
des produits enfin c'est super
intéressant la vision qu'il a de
agriculture il teste plein plein de
choses à grande échelle avec je pense
qu'on a des dizaines de milliers d'
analyse de sève sur toutes les cultures
tout être humain quand il progresse vite
aussi qu'il faut savoir minimum de suivi
à savoir à quelle heure on améliorer
c'est important parce que c'est nous
permettent de revenir puis moi ça m'a
permis de comprendre qui traitait une
mère de l'après midi
at elle des fois on est obligé mais
j'ai pas moi sur un soin un passage du
glyphosate pour contrôler un couvert
végétal
côte d'ivoire je comprenais pas au début
on a tu piques lausanne et qui coupait
on a je sais pas quoi avec les deux
piasses à drake et puis en fait il le
passait à midi puis à partir du moment
où on a dit ion
s'organisent autrement et à 9 heures du
matin c'est une fini d'être passé
là les doses elles ont été divisées par
2 quoi
et sur le ph de l'eau pour des shows
enfin je sais pas car il ya plein de
choses comme ça le ph de l'eau sur lui
des choses assez
on
peut vite baissé les doses très
fortement on reprend en parlant du
glyphosate aussi des inconvénients
c'est ni tout noir ni tout blanc
bien utilisé c'est c'est intéressant
mais on sait qu'il ya des
problèmes aussi
entre autres c'est un créateur du
manganèse
bon ça je sais pas mais bon rousse à
quatre heures du mans donné il va
bloquer le manganèse
donc si c'est pour contrôler un gros
couvert végétal quart équilibré le sol
très bien c'est pour faire comme ans
comme en argentine ou dans le sud du
brésil c'est une aberration totale
c'est là l'utiliser enroulé secours ou
en lac en temps c'est intéressant
utilisé comme comme moteur du système
on va dans le mur quoi
l'annoncé s'est il faudrait demander
stefan et reining des stands ont là j'ai
du mal à tout suivre de près est-ce qui
se pressent là j'avais passé une journée
à les identifier
chaque emplacement de mesures pour être
sûr que la stagiaire soit le fasse comme
il faut donc dans la journée on a
couvert la moitié du var
maintenant que chez moi
oui là c'est oui c'est juste sur une
fois il faut il faut qu'on suive ça en
fait ça au départ c'étaient des mesures
qui sont faites pour calibrer pour
calibrer on a besoin de diversité donc
on dit en même temps commence par en
profite pour commencer à prendre un peu
d'infos sur sur les fêtes différentes
pratiques
c'est vraiment par les scanners qu'on va
pouvoir qu'on va pouvoir structurer ça
comme il faut et à voir les répétitions
et
savoir ce qui s'est passé exactement
vidéo
chez cependant
à cette date de quand
je joue sur la
bbc sur la berline plus tôt
c'est peut-être sur la belle vie mais
plutôt parce que vers de ta je suis
assez souvent ce qu'il n'y a pas eu
grand chose de nouveau là dessus on sait
doit être sur la belle vigneault et
abbas se traduit très bien oui cbc c'est
possible ouais c'est possible
plutôt sur la belle vie il faut que
j'aille regarder ben justement
voilà alors dans la même série on a
du cinsault en bleu du cinsault avec des
engrais lala ce que j'ai mis le ph et le
red box et la gemmi la conductivité
électrique en fonction du pe plus php
comme ça avec ça on a à peu près toutes
les infos la bioélectronique qui vont
donc du cinsault du cinsault avec de
l'engrais du cinsault avec du fumier et
du fumier plus engrais donc ce qu'on
voit en gros fumier fumier puissant gré
on arrive alors ça a évolué comme ça et
ça celui-là il tourne comme ça ça paraît
dans le sens de la flèche avec 19
juillet 2 août 18 août et 31 août
donc là on est dans l'eau dans le temps
comment ça bouge dans le temps ok donc
en gros parce qu'on voit quand même
c'est que le fumier
c'est le gris et john sano
sanou baisse surtout en pe plus ph il a
fait en général je mêle on le voit ici
là le pe plus ph il est là donc c'est
surtout ça nous montre la conductivité
près là ça bouge dans tous les sens non
on commence à distinguer des trucs sa
part à peu près du même endroit il a
évolué différemment
sa
vie mais c'est les premières mesures il
va falloir comprendre ce qui se passe
qu'est ce qu'elle fait dans cette
parcelle c'était comme ça
quand
la mesure pour la mesure et c'est aussi
pour
là c'était pour paramètres et beaucoup
mais en même temps ça commence à donner
des infos quoi justement à ça ça va
poser des questions pourquoi pourquoi
des fois ça revient dès fois ça passe
qu'est ce qui fait que l'engrais fait
chuter la
par contre lang lang rays convoi en
conductivité on est bien plus bas c'est
ce qui est assez surprenant
a priori des engrais chimiques enfin
c'est mignon comme ça ça augmente la
conductivité
alors pas ou peut-être que ces noms
c'est peut-être le côté nitrates ou en
charge en eau
oui c'est fait c'est peut-être ça c'est
peut-être ça c'est peut-être un côté
nitrates il va falloir qu'on regarde
quel ongrais qu'elle avait pré condition
est que
la conductivité des micro organismes
de plus on a mis organisme
au sein d'accor
dans la plante ou dans le sol
là on est dans la plante donc
je pense ça doit être dans le sol ouais
ouais oui ce qui est logique way
l'âge l'âge des cépages là c'est
celui-là il est intéressant c'est qu'on
a en bleu 1st thought 15 ans en orange
un grenache de 6 ans et en gris un
grenache de 40 ans et donc en gros le
cinsault de 15 ans et le grenache de 40
ans sont assez proches et le grenache de
6 ans il est bien plus oxydé
il un ph qui bouge beaucoup plus
j'ai demandé aussi à ce que les
hypothèses ça peut être que une plante
quand elle est jeune elle est peut-être
pas encore complètement adaptée à son
milieu au sens où elle a peut-être pas
mobilisés sont gênantes tous cas là je
pense que c'est un truc dans le genre
s'adapter complètement et du coup et
puis à la moindre réserve
enfin il ya toute une gestuelle
voilà elle est plus sensible au stress
hydrique enfin mais je pense ça c'est
quand même intéressant comme comme
premier
je pense que normalement oui on était
sur le sait la même seul mais c'est le
même coin et puis c'est marrant c'est
quand on peut plus ph on voit qu'on est
bien plus haut quoi c'est pas une
histoire de conductivité c'est vraiment
du pe cus ph
voilà pour après pour comprendre
pourquoi certaines vignes prennent du
mildiou et pas d'autres éléments déjà
l'âge là dedans ça nous donne un peu
comme ça c'est plus parlant oui oui bah
c'est après c'est bien d'avoir les deux
formes parce que là on rajoute la
conductivité alors même si pour
l'instant on sait pas trop bien
l'utiliser il ya des il ya quand même
des infos qui nous disent qu'il faut
qu'on la mette et quand il faut
justement il va falloir acquérir toutes
ces données pour mieux comprendre
comment ça se passe
oui il ya une variabilité dans les
mesures c'est pour ça que je pense qu'il
faudra faire une dizaine de mesures par
par traitement ce qui était quasiment
impossible à faire
avec ça et ce qui va prendre ce qui va
prendre deux minutes par traitement avec
les scanners
voilà pour pour faire ça c'est voilà ça
c'est chaque chaque jour c'est une
journée de mesure de sa doit etre il
doit y avoir la moyenne de quatre là il
faudrait que mettre les écarts types aux
2/3 de 3 pourrait commettre les écarter
mais j'ai pas miser quartier parce que
j'attends d'avoir les résultats des
scanners pour avoir si ces répétitions
lieu de trois voilà pour l'instant on a
fait trois mesures en électrochimie et
puis 3 scan qui correspondent plus trois
autres scan
donc on
rétroactivement maintenant qu'on a
calibré en double la puissance
statistique donc il faut qu' il faut que
je fasse les analyses mais j'y arrive
pas tout faire
voilà mais ça je suis là il est
intéressant
enherbement alors voilà là c'est
comprenait pas trop il respecte le mans
justement des précisions donc en herbe
et se met en fête c'est un rang sur deux
qui enfin rang
sur deux en herbe et semer un rang sur
de travail et
naturel sol nu c'est un rang sur le sol
du travail et un enfant sur de
végétation naturelle et naturel plus
mais c'est un grand naturel un an un
rang en herbe est
d'accord
e
c'est comme les dates ouais je vois j'ai
fait ça vite mais c'est du même ordre de
grandeur assez la baisse et la même
stagiaire c'est en gros de début juillet
à début septembre
annoncé la sela vigne c'est la vie c'est
la fin et c'est la feuille
c3 à ses proches chaque point un point
là c'est la moyenne de 3
pour l'instant mais j'ai espoir on va
avoir de quoi faire la moyenne de 6 donc
ce sera le seul ennemi depuis l âge que
c'est lui est ce que la fit ni el para
ps3 et que si elle s'assit dif ils vont
avoir du mal aussi quand même
la question c'est c'est ce que
est-ce que ça le cas lisez trop sait pas
ce qui nous amène au mildiou
si on s'oxygène plus quoi
parce qu'a priori le mildiou c'est quand
même un ami seth qui est dans des ph
bien plus élevé donc ça a été fait sur
les chaussures le vernis votre feuille
aussi c'est parce que le fils fait dix
enfants et jeunes et on prend toujours
là une feuille en pleine activité
photosynthétique
dans les dents les plus jeunes feuilles
en pleine activité photosynthétique et
qui soit à la lumière si on va en
prendre une qui est derrière et cachés
dessous
ça va pas faire la même chose ce qui est
sale et l'idée c'est de
il vous dit il n'y a pas les répétitions
statistiques c'est vous donner une idée
de ce qu'on va pouvoir en faire de
maître de valada voir les répétitions
qu'avec les scanner on va pouvoir le
faire ces mesures au départ c'était
surtout pour calibrer
mais on
calibré pour calibrer c'est un peu
débile autant commencer et puis surtout
ça amène de la diversité dans ce conte
calibre parce que si on call him to avec
les mesures dans la même parcelle et ben
on sera super pointu sur cette parcelle
là mais dès qu'on sortira on sera dans
les choux complet donc là il ya une
diversité de parcelles et tout voilà au
niveau conductivité en plus donc par
contre là ce qu'on voit surtout c'est
que ambiance
alkali nisand le pe plus péage qui
montrent une mante fort quoi
j'ai mis la flèche dans le sens où ça va
ben oui normalement oui c'est réducteur
oui il ya
eu enfin c'est quand même le naturel sur
le nuc est le plus oxydé tout le temps
quoi
ouais elle aura près là il faut voir la
puissance statistique les écarts types
qui aurait eu ma chance et pas forcément
des différences significatives l'on
n'est quand même pas sur des gros écarts
donc il faut cesser à prendre avec
précaution là c'est pour voilà pour vous
donner une idée de souvenirs qu'on peut
aller
potentiel qu'il ya derrière quoi après
ça on appris c'est pas c'est pas publier
c'est pas publiables dans l'état
peut-être qu'avec les six répétition va
pouvoir et safran beaucoup de temps donc
on va essayer de comprendre mieux parce
que doivent s'y vu tout ce qu'on a à
prendre voilà si
sur le riz pourquoi sa selle kelly nice
sur ce qu'on a en france est-ce que
c'est l'eau et c'est un machin il va
falloir le travailler avant de publier
en disant on sait pas quoi
donc voilà
je n'ai pas mis sur le million parce que
c'était trop trop trop variabilité mais
en gros l'idée c'est de suivre les
parcelles où on voit les milieux qui
arrive
c'est ça c'est comme ça qu'on va y
arriver c'est faire des mesures
pétition en plus vous connaissez
dans vos parcelles il ya des coins qui
prennent plus le mildiou d'autres
les creux les enfin ces machins là
ça sera de suivre très régulièrement
dans ces parcelles l'alleud
you et puis voir l'apparition des
premiers
netbooks sont ignorés de police en tome
car qu'apparaissent quoi sachant que
c'est l'oeuf et la poule quand on sait
que ça paraît parce que c'est trop oxydé
et que la réaction c'est de sur oxyde et
donc il faut le chopper rapidement pour
avoir son niveau de départ donc ça c'est
du suivi régulier où il faut voir si on
peut pas arriver enfin s'il n'y a pas
des gens qui arrivent à le cultiver à
peu près sûr au moins sur des feuilles
enfin quand
c'est nous on n'est pas à l'abri de ça
c'est
si mais c'est pour ça qu'il va falloir
structuré comme il faut pour
le plus dingo sais que vous allez avoir
des trucs une année pas l'autre mais ces
voix là mais
mais oui mais ça va bouger différemment
ça pour me permettre de comprendre que
c'est l'effet de ce stress là et que
je
sente spectraux ces mêmes on en
parlerait même pas si je n'avais pas le
pétrole how did on veut mesurer je vous
dirai pas non attendez pas ce que parce
que je vous dit vous allez passer une
journée pour faire
la langue graphique comme ça ces quatre
journées complètes
ok
donc donc vous allez dire non on va pas
on va pas le faire avec le spectre au un
graphique comme ça c'est
c'est quoi c'est que ces quatre fois une
heure quoi
donc c'est pas ça change
ça change la donne et même pas c'est
même pas quatre fois une heure s'il ya
trois noms il ya trois points là c'est
même pas ces
trois points à chaque c'est le temps
c'est de passer d'une parcelle à une
autre
ces trois fois dix minutes là et enfin
quatre fois dix minutes compétition et
le vent de 2500 euros je crois
après si air plein de mesures en
parallèle je pense qu'il ne sera pas au
même prix toi parce que tu aides à
calibrer donc voilà mais
moi ça c'est d'un premier prototype
c'est même le premier prototype ça doit
être exemplaire un
100m ils ont même mis mon nom dessus
sont chanson était maintenant en fait il
ya un rail ici qui permet de glisser une
cuve qui permet de faire les liquides
ayana spécial liquide aussi ouais
vous élever un peu claires où elle et
des vins blancs
et l'ont fait en ans a évolué en suisse
ont posé la question est ce qu'on y va
est ce qu'on n'y va pas s qui
commercialise la baïse allez on y va
sachant que la prédiction sur la vigne
pour l'instant et encore enfin pour ici
ça doit être pas mal du tout
ce calibre est pas loin d'ici quand même
donc
voila voila donc
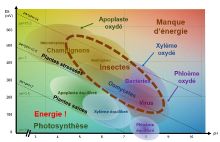
si on finit sur la plante donc en gros
c'est la plante c'est contrôlé par des
signaux ph redox partout dans tous les
sens et niveau ph credoc de la plante il
dépend du raid du sol et ont toutes la
photosynthèse et un peu de la
respiration aussi la nuit ça s'oxyde
parce que ça respire
et le niveau réel de la plante il dépend
de l'organe de sa position sur la plante
de lâches
du génotype espèces et variétés donc il
faut prendre tout ça en compte pour
comprendre derrière comment ça ce
comment ça se passe
Portail thématique





